the assumption of responsibility for the welfare of the world
|
STEWARDSHIP the assumption of responsibility for the welfare of the world |
|
SITE MAP
|
REFERENCE BASE
Life is a process in which systemic entities sustain themselves, or specifically maintain their internal processes, by drawing resources from their environments. But there is no empirical definition that completely identifies what life is and is not in common human usage; humans, even human scientists, generally identify life heuristically, with the term life being reserved for processes sufficiently similar to human life, while also not primarily the product of human technology thus, life involves entities made of cells, based on proteins, nucleic acids, and water. As such, the term excludes phenomena such as storms, as well as artificial machines. At the same time, the prospect of extraterrestrial life is accepted even though the exact structure could be significantly different. The cell is the basic unit of Earth-based life, consisting of genetic material in a water-based environment, contained within a membrane. An organism is a cell or group of cells functioning as an independent, integral system, capable, in its proper habitat, of sustaining itself, and generally reproducing at some point in its life cycle. Organisms are divided into eukaryotes, those that have eukaryotic cells (from Greek ΕΥ- « eu- », good + ΚΑΡΥ- « karu- », kernel, nucleus), and prokaryotes, which consist of a single prokaryotic cell (from Greek ΠΡΟ- « pro- », before + ΚΑΡΥ- « karu- »). Eukaryotic cells contain organelles, which are membrane-enclosed compartments, one of which, the nucleus, contains the cells genetic material. Prokaryotic cells do not have organelles. Eukaryotic cells evolved as symbiotic systems of prokaryotes, several single-celled organisms living inside another single-celled organism. Evolution A species is a breeding population a group of organisms that reproduce with each other, and produce offspring who themselves can reproduce. The reproduction establishes continuity over time, so that the species continues to exist even when earlier members have all died. In most uses, organisms are part of the same species not when they merely can reproduce with each other, but when they do reproduce with each other. But the identification of species is also somewhat heuristic, since populations can be sundered, evidence of reproduction is not always available, and not all individuals of any species will or can actually reproduce. An ecological niche is a food or energy source or a habitat that is available to be exploited, a function in the ecosystem available to be filled. A niche is basically an opportunity. Every species can be said to fill an ecological niche, and every species depends for its survival on its suitability to a particular environment. This, in turn, requires adaptation to that environment through the process of evolution. Biological evolution is the gradual change of species over time, through random mutation and natural selection. Each organism develops according to its genes, its genetic code, stored in molecules of deoxyribonucleic acid (DNA) on chromosomes in each cell. A mutation is a change in that genetic code; it is an accidental corruption that gets introduced to the code through imperfect reproduction, leading to slight differences between certain individual organisms and their offspring. But these mutations serve an important purpose. Given enough mutations, eventually a mutation will emerge that gives its individual organism a competitive advantage that allows that organism to fill a new ecological niche, that gives it a better chance than the rest of its species at surviving long enough to reproduce. If so, that mutated gene will be passed on to offspring, who will also have the competitive advantage. Over time, natural selection occurs. In natural selection, or survival of the fittest, whichever individual organisms are best adapted (through random mutation) to compete for necessary resources in a particular environment will ultimately survive and reproduce in that environment, while other organisms will not. Parallel evolution is when features or functions evolve separately in two or more different lineages. For example, birds and bats both have wings, not because they are descended from a common ancestor with wings (they are not), but because the feature developed independently. The concept of parallel evolution is important in understanding ecorealms (see below). Many different plant lines evolved a tree form, because there is a common ecological niche for a tall plant that can compete for solar energy, and a sturdy (woody) trunk is an adaptation that allows a plant to grow to this kind of height. The famous question Which came first: the chicken or the egg? is meant to be a paradoxical puzzle, but in the theory of evolution, there is a definitive answer: the egg came first. Every chicken hatched from a chicken egg. Not every chicken egg was laid by a chicken, though. At some point, there was a chicken egg fertilized and laid by two birds who were not quite chickens very, very close, but not quite. Through a random mutation, the egg received DNA that was ever so slightly altered, and ended up as the DNA of a modern chicken, rather than the almost-chickens that were its parents. Classification Traditionally, organisms are classified hierarchically. All organisms are assigned to a kingdom, and then to a phylum or division, a class, an order, a family, a genus, and a species. Each order, for example, belongs to a class, and contains one or more families. These levels are named somewhat arbitrarily; most of the names simply mean kind. Collectively, these levels are called taxa, the plural of taxon, which also means kind. (Taxon is actually a Neo-Greek coinage from taxonomy, itself from a later French coinage from Greek ΤΑΞΙΣ « taksis », arrangement, ordering + ΝΟΜ- « nom- », law.) Humans are classified thus:
In short, modern humans are Homo sapiens sapiens, or simply Homo sapiens. There have been other species in our genus, such as Homo erectus, but they are all now extinct. Taxa are conventionally named in (New) Latin, and cited as plural nouns, except genus and species, which are a singular noun and an adjective, respectively. The English versions of our taxonomic names are animals, chordates, vertebrates, mammals, primates, and hominids, while Homo means human, and sapiens means knowing. In the traditional taxonomy, birds (Aves) and reptiles (Reptilia) are both classes. Birds, though, are now recognized as dinosaurs (Dinosauria), which were already recognized as reptiles and it is impossible, in the traditional taxonomic system, for one class to be a part of another class. The solution to this problem has been the development of cladistics. A clade (from Greek ΚΛΑΔΟΣ « klados », branch) is a group of organisms with a common ancestor. One advantage of cladistics over traditional taxonomy is that new relationships for clades like Aves do not require a wholesale revision of the system. A second advantage is that it eliminates the impression that all taxa at a particular level are somehow equivalent, so that all families are presumed to represent the same number of species or the same number of individuals. Finally, cladistics eliminates the need for unnecessary intermediate taxa as well as the need for interspersed taxa (supertaxa and subtaxa like the subphylum Vertebrata); a particular lineage can have as many or as few clades as is appropriate.
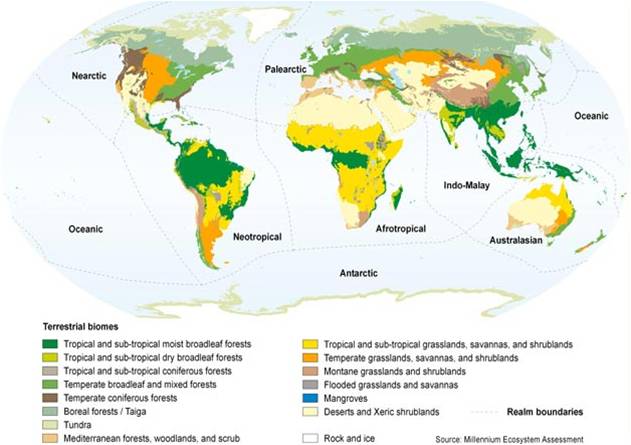
Ecology is the law of interaction and coexistence of living things, or the study of how they do so (from Greek ΟΙΚ- « oik- », household + ΛΟΓ- « log- », account, study). The premise of ecology is that life exists interdependently in systems. Ecology is closely connected to biogeography, the distribution of living things, or the study of that distribution. A biocommunity is the collection of all organisms living together in a particular place. An ecosystem is a biocommunity together with all the non-living elements of the environment water, soil, rocks, climate, etc. A bioformation is an area of similar vegetation; it is the equivalent of a biocommunity or ecosystem, but only concerned with plants, particularly the dominant plants and their physical configuration (tall, short, high canopy, undergrowth, etc.). A biome is a kind of ecosystem. A biostructure is a kind of bioformation. Accordingly, we can say that a given ecosystem is a local example of a biome, and a given bioformation is a local example of a biostructure. The map at left shows a particular interpretation of world biomes. Thus, rainforest is a biome; the Amazon rainforest is an ecosystem. Similarly, rainforest is a biostructure, and the Amazon rainforest is a bioformation. The Amazon is a bioformation when we are just considering that it is a local collection of a trees in a humid environment; it is an ecosystem when we consider all of the organisms and all the elements of the environment. As rainforest is a biome and a biostructure, forest is also a biome and a biostructure, merely more general. We could also speak of a more specific biome and biostructure, such as tropical rainforest. We can speak of other forest biomes, like deciduous forest, which can also be further specified. (Note: within ecological contexts, biocommunites, biostructures, and bioformations are simply called communities, structures, and formations.)
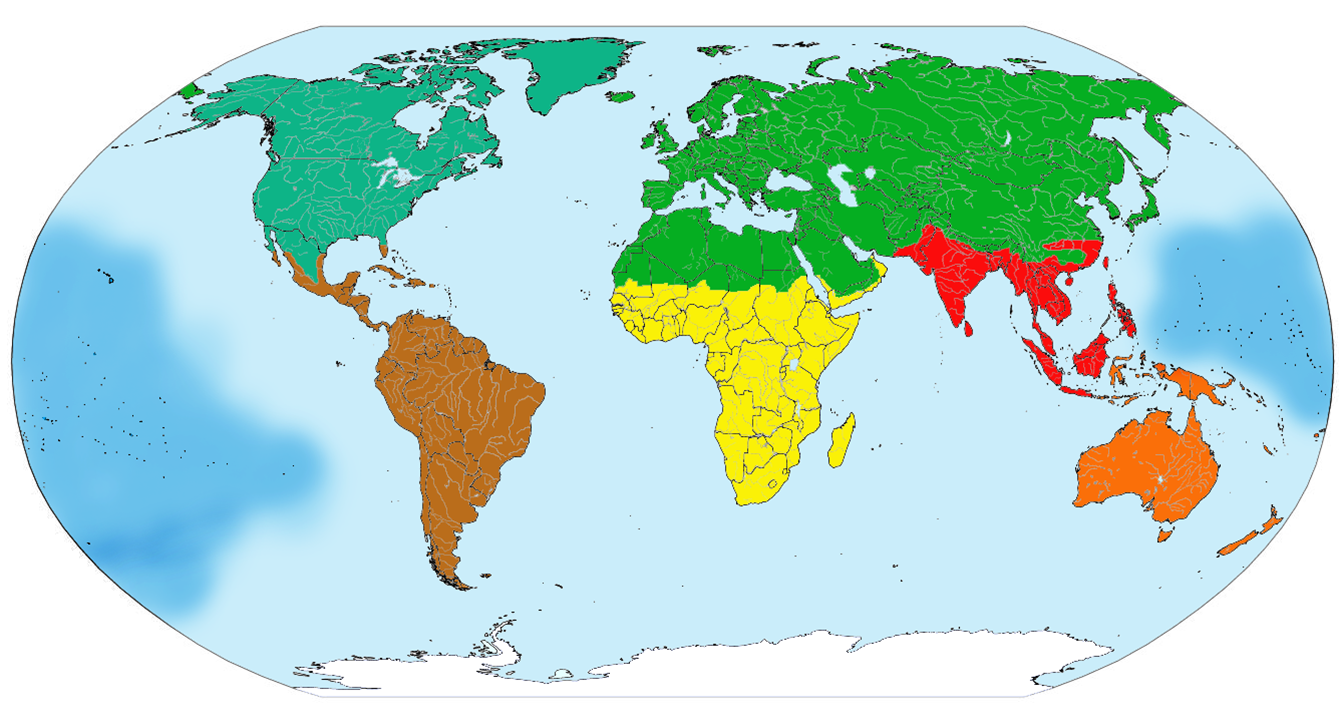
All of the terms above describe entities without clear borders. Instead, we should expect transition zones, where one area blends into another. At best, we can define an ecosystem by a particular feature. If we use a species, for example, we can then locate the limits of that species; but the species will not generally cease suddenly along a line. Rather, it will become less and less prevalent until there are just a few individuals living surrounded by other species that we associate with completely-different ecosystems. These places biocommunities, ecosystems, and bioformations are, then, best understood as defined in specific ways for specific purposes. An ecorealm is a major division of the world based on relatedness of species. Plants and animals in one ecorealm are more closely related to other plants or animals in the same ecorealm than they are to similar plants or animals in other ecorealms. For example, trees in the Afrotropical ecorealm are more closely related to shrubs in the Afrotropical ecorealm than they are to trees in other ecorealms. Ecorealms exist primarily because of barriers to species migration the ocean, the polar climate, the Sahara desert.
The ecorealms depicted are: Nearctic (blue-green); Neotropical (brown); Palearctic (green); Afrotropical (yellow); Indo-Malay (red); Australasian (orange); Oceanic (medium blue, covering the Pacific islands); Antarctic (white). These are the ecorealms used for all organisms; if instead we were just considering plants, or just animals, wed have a different (though similar) set of ecorealms. And the exact location of these ecorealms is, like many things in science, still being debated and worked out. Alfred Russel Wallace (who was also famous for independently formulating the theory of evolution, along with Charles Darwin) began this branch of biogeography by identifying the line between the Indo-Malay and Australasian ecorealms (as they are now called); this was subsequently called the Wallace Line. But other researchers have proposed that the boundary between the ecorealms lies along a different line. This doesnt change the basic idea, though; they agree in the concept, and are just working out the details. (The prefixes ne-, from Greek ΝΕ- « ne- », new, and pale-, from ΠΑΛΑΙ- « palai- », old, are simply references to the New World and the Old World, while -arctic and -tropical in this context mean towards the Arctic and towards the tropics.)
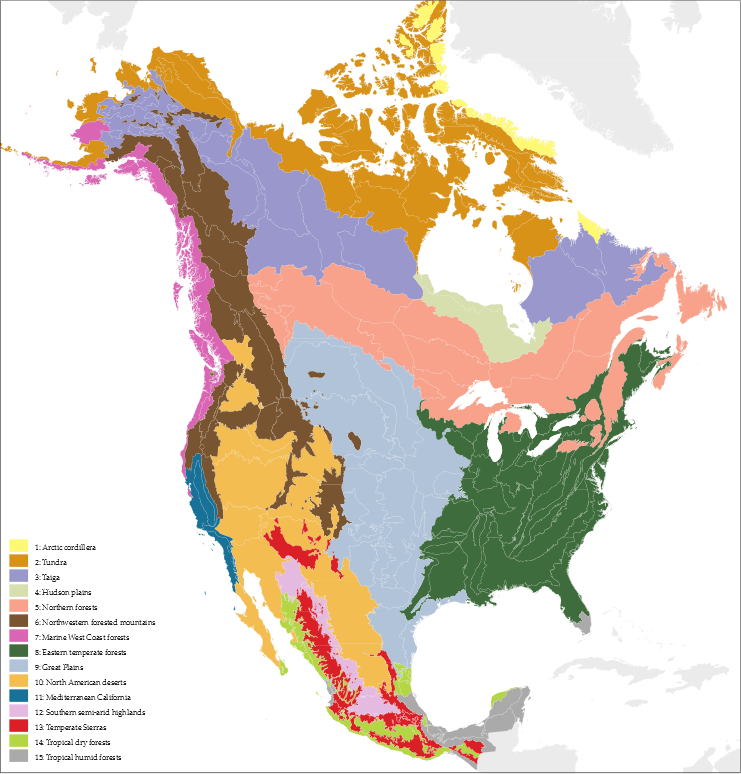
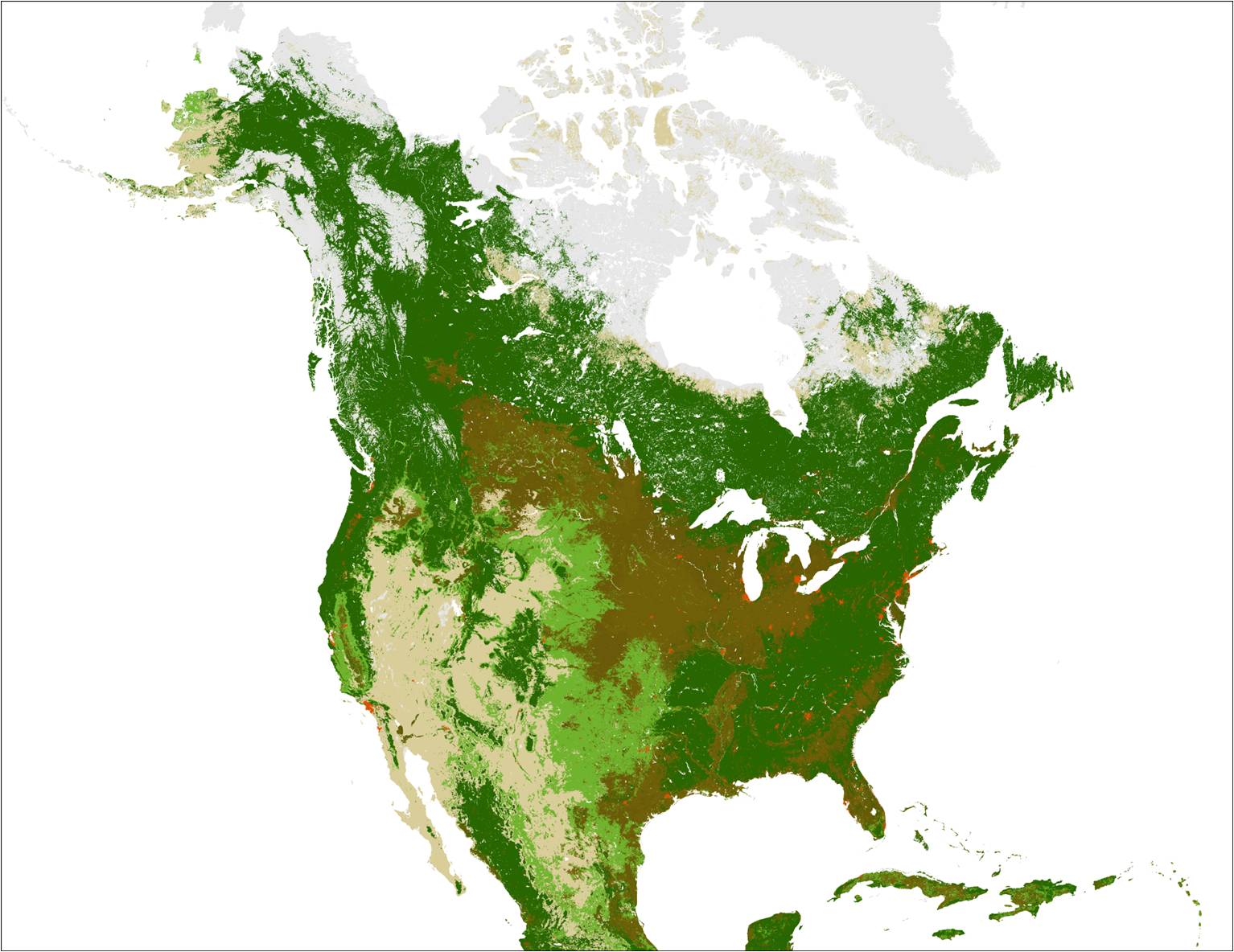
Note the difference between the center of North America in this map (right) versus the CEC map above. The Great Plains, the light blue ecoregion in the CEC map, is colored as two different land covers. The light green is the grassland of the original Great Plains. The dark green is forest. The dark brown is farmland, which doesnt appear anywhere on the CEC Level I map, despite the prevalence of agricultural land in much of North America. And yet it is obviously an important empirical feature of our environment. The dominant plant life in most of the brown area is corn, wheat, soybeans, or another food crop. Energy cycle Photosynthesis is the process where certain cells, photosynthetic cells, convert solar energy into stored chemical energy. Photosynthetic cells are found in plants, but also in various algae. The basic chemical equation for photosynthesis is: 6 H2O + 6 CO2 + energy → C6H12O6 + 6 O2 Water (H2O) and carbon dioxide (CO2), with an input of energy from the sun, yield glucose (C6H12O6) and oxygen (O2). Glucose is a simple carbohydrate, and a base form of energy used in cellular respiration, the metabolism of chemical energy that allows living cells to function. The basic chemical equation for cellular respiration is the reverse of photosynthesis: C6H12O6 + 6 O2 → 6 H2O + 6 CO2 + energy Glucose and oxygen are consumed, energy is freed up, and water and carbon dioxide are produced. All cells, including plant cells, respire. Thus, instead of viewing plants as making oxygen and carbohydrates for animals, the correct view is of plants photosynthetic cells as making oxygen and carbohydrates for the plants themselves and non-photosynthetic organisms. We generally speak of four trophic levels (from Greek ΤΡΟΦΗ « trop‛ē », food). The lowest level is the producers, plants and other photosynthesizers, those organisms that convert solar energy into usable chemical energy. Primary consumers are the organisms that eat producers mostly, in other words, herbivores. Secondary consumers eat primary consumers, which accounts for most carnivores. Tertiary consumers eat secondary consumers; many tertiary consumers are scavengers (that is, animals who eat animals after they die, rather than by killing them). At each trophic level, 90% of available energy is consumed in cellular respiration, leaving only 10% of that energy to feed the next level. Primary consumers are only getting 10% of the energy consumed by the producers; secondary consumers are only getting 1% of the energy consumed by the producers. Energy is not being wasted. Trophic level does, however, affect resources used for a given amount of energy. One argument for humans to eat at a lower trophic level is the reduction of the amount of land, water, and other resources needed to produce our food. If we ate plants directly instead of eating animals that eat plants, we would need only a tenth of the resources to get the same amount of energy, leaving the rest for other uses, leaving more land as wilderness, etc. There is nothing inherently good or bad in nature or an organisms place on the trophic scale, even a human organism; as humans we interject such judgements based on our values, including what is good for us personally or as a species, which generally includes the preservation of our own environment. Disturbance We can define an equilibrium as a persistent state in a system resulting from a stable balance of forces. A disturbance temporarily or permanently upsets an equilibrium. Disturbances things like fires, earthquakes, hurricanes and tornadoes, severe thunderstorms are natural and normal, and ecosystems are adapted to certain disturbances. What matters are time scale and severity; ecosystems adapt to disturbances that happen with a certain frequency and a certain intensity, but anything more could permanently upset an equilibrium. Again, disturbance is not good or bad per se, but human-caused disturbances raise the prospect of permanently upsetting an equilibrium we ourselves rely on.
© O.T. FORD |
THE STEWARDSHIP Home of the Stewardship Project and O.T. Ford |